


01/03/2010
L'Europe est un tout unique
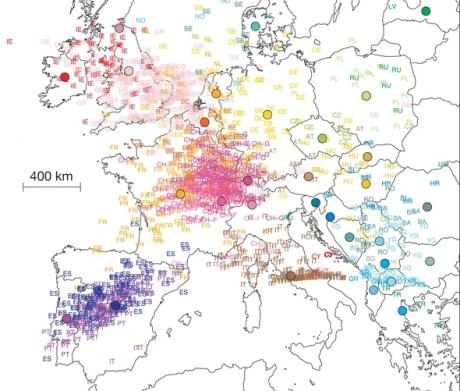
| GENETIQUE – Le profil génétique d’un Européen permet de savoir de quelle région il vient… En faisant analyser vos gènes, on pourra bientôt savoir si vos aïeux venaient de Picardie, de Bavière ou de Lituanie. C’est en tout cas ce que promettent les résultats de deux récentes études de génétique des populations en utilisant une nouvelle technique d’exploration du génome humain permise par les puces ADN : l’analyse des mutations individuelles (SNP). Des profils et des distances John Novembre et une équipe americano-suisse ont analysé une large base de données répertoriant les 200 000 variations génétiques individuelles d’un échantillon de 1387 Européens vivant dans 36 pays. A l’aide d’outils statistiques, ils ont ensuite établi un profil génétique numérique pour chacun des sujets, ce qui leur a permis de déduire une distance génétique entre chaque sujet. Géopolitique des mutations Ils se sont ainsi aperçus que la distance génétique entre deux individus était à peu près proportionnelle à la distance entre les régions où ils vivaient. En projetant graphiquement ce résultat, ils ont ainsi constaté que la carte des profils génétiques recouvrait la carte géopolitique de l’Europe. Ces travaux ont été publiés dimanche dans la revue « Nature » ; les résultats rejoignent d’ailleurs ceux déjà obtenus dans une étude similaire publiée début août dans la revue « Current Biology » et dirigée par le généticien hollandais Manfred Kayser. Généalogie génétique Les deux études montrent qu’en dépit des échanges incessants, des migrations et des invasions, les Européens gardent une odeur de terroir dans leurs gènes. « Cela rend envisageable la mise au point de tests génétiques permettant de connaître les origines géographiques d’un individu » explique John Novembre à 20Minutes.fr « sur notre carte géo- génétique, d’après leur profil, 50% des individus se retrouvaient à moins de 310 km du centre de leur pays d’origine, et 90% à moins de 700 km ». Pas de races européennes Toutefois, le chercheur tient à préciser que cela n’implique pas l’existence de « sous-races » européennes génétiquement différenciées. « En moyenne, les différences génétiques entre populations européennes sont négligeables : sur une échelle de 0 à 1 (1 représentant deux génomes complètement différents) le degré de différentiation génétique est d’à peine 0,004 entre régions européennes ; absolument pas de quoi parler de sous-races, en tous cas au sens biologique » poursuit John Novembre « la possibilité de déterminer la population d’origine de chaque sujet n’a ici été possible qu’en prenant en considération les dizaines de milliers des mutations ponctuelles dont il était porteur ; on a ensuite repéré celles qui étaient les plus fréquentes dans les différentes populations européennes pour inférer l’origine géographique de chaque individu ». Yaroslav Pigenet |
23:43 Publié dans Dépêches | Lien permanent | Commentaires (25) | Tags : génétique, europe-nation, adn, europe-patrie |
Commentaires
Écrit par : Anton Cusa | 11/07/2014
Écrit par : Anton Cusa | 11/07/2014
Écrit par : anton Cusa | 11/07/2014
Quant à l'origine que vous donnez des IE, elle n'est pas compatible avec les données linguistiques, les seuls fiables en la matière, ni avec la mythologie comparée (et surtout avec sa symbolique associée).
Au-delà de la subdivision au sein de l'espèce en quatre sous-ensembles puis des adaptations in situ au sein de chaque ensemble, on ne peut rien déduire.
Vous n'avez pas tort sur le processus initial mais sur la chronologie. Ce changement d'échelle change tout.
Enfin, vous passez sous silence la forte homogénéité des populations européennes que cet article évoque, en dépit des quelques haplotypes "transphyliques" que vous évoquez remontant sans doute à une époque bien plus ancienne (probablement avant qu'homo erectus ne se divise en branches).
Cordialement.
Écrit par : Thomas FERRIER | 11/07/2014
Écrit par : Thomas FERRIER | 11/07/2014
Écrit par : Anton Cusa | 12/07/2014
Écrit par : anton Cusa | 12/07/2014
Pour R1b, voir notamment : "Ancient History of the Arbins, Bearers of Haplogroup R1b, from Central Asia to Europe, 16,000 to 1500 Years before Present, de Anatole A. Klyosov"
http://www.scirp.org/journal/PaperInformation.aspx?paperID=19567#.U8FEApg0uIg
Écrit par : Anton Cusa | 12/07/2014
Écrit par : anton Cusa | 12/07/2014
Que les Néanderthaliens et les hommes européens modernes présentent des caractéristiques morphotypales extrêmement proches est également une donnée importante. Même si les uns ne descendent pas des autres, mais sont deux évolutions de l'homo antecessor, l'évolution proprement européenne d'homo erectus.
La théorie de l'évolution multirégionale, qui n'interdit pas des flux géniques à la marge (pouvant largement expliquer ce que vous évoquez), repose avant tout sur des données anthropologiques et archéologiques.
Considérer que l'ancêtre commun aurait 140.000 ans et viendrait d'Afrique est une théorie, fortement idéologisée, reposant sur une datation basse des données génétiques. Or, si ancêtre commun incontestable il y a, c'est homo erectus.
Écrit par : Thomas FERRIER | 12/07/2014
Écrit par : Thomas FERRIER | 12/07/2014
Écrit par : Anton Cusa | 12/07/2014
- la confusion entre flux génique et ascendance commune
(la notion d' "ancêtres communs" est-elle même un présupposé d'adhésion à la théorie monophylétiste)
- une datation qui repose là aussi sur le même présupposé
Qu'un flux génique ait pu introduire des gènes qu'on retrouve prioritairement ailleurs au sein de la population européenne, sans que cela ne remette en cause son homogénéité (nécessairement relative), personne ne le conteste.
Enfin, il faut prendre en compte l'ensemble des gènes, ce que cette étude fait, et celle-ci implique une évidence, l'origine commune de toutes les populations europoïdes.
Écrit par : Thomas FERRIER | 12/07/2014
Écrit par : Thomas FERRIER | 12/07/2014
Écrit par : Anton Cusa | 12/07/2014
Écrit par : Anton Cusa | 12/07/2014
Écrit par : Anton Cusa | 12/07/2014
L'espèce humaine n'est pas différente des autres espèces mammifères, à savoir une subdivision ancienne en plusieurs sous-espèces. Ce qui implique une origine commune à tous les membres d'une même sous-espèce. Quant à l'interfécondité, elle est établie entre le lion d'Afrique et le lion d'Asie, par exemple. Ce qui ne signifie pas une différenciation récente.
J'ai affirmé que l'homo sapiens europaeus descendait comme l'homo neanderthalis d'un ancêtre commun. Je n'ai pas affirmé interfécondité entre les deux. Je pense plutôt à une variante "aberrante" hyper-spécialisée de l'un dans un environnement spécifique. En clair, les néanderthaliens se seraient rapidement isolés et différenciés de l'autre branche cousine.
La seule explication reste un flux génique constant, pas forcément important néanmoins, ce qui est admis par les partisans de la théorie de l'évolution multirégionale.
Quant à l'interprétation des données, qui est au moins aussi importante que les données elles-mêmes, elle repose souvent sur des "postulats de principe", comme dans la fameuse affaire Clovis, où les chercheurs refusaient de fouiller en dessous de -12.000 ans, alors qu'on a fini par trouver la preuve d'un peuplement plus ancien de l'Amérique (sans doute solutréen).
Anthropologues et généticiens "médiatiques" s'opposent. J'ai évoqué mes propres intuitions avec des chercheurs en ce domaine, qui ont reconnu la faiblesse du discours sur les fameux "haplotypes" et aussi sur leur caractère de marqueur d'ascendance.
Quant à savoir si cela est dominant, visiblement c'est moins de 0,004% en moyenne.
Cet article donne de l'importance à la notion d'homogénéité donc de profonde parenté, ce qui ne signifie pas uniformité. Il n'est en rien remis en cause.
Écrit par : Thomas FERRIER | 12/07/2014
Je n'ai ceci dit pas besoin de m'appuyer sur des données génétiques. Je trouve simplement que cet article était intéressant car rappelant une évidence.
Écrit par : Thomas FERRIER | 12/07/2014
Écrit par : Anton Cusa | 13/07/2014
Quant au peuplement néolithique de l'Europe via le moyen-orient pré-afrasien, c'est une hypothèse effectivement majoritaire et assez crédible, mais il ne faudrait pas négliger l'idée que le moyen-orient a pu préalablement être peuplé de populations venues d'Europe, dont les ancêtres des Sumériens et des Hourrites.
Un certain nombre de données laisse penser que le peuplement de l'Europe par l'homme moderne se soit fait de l'est vers l'ouest, mais pas du sud-est vers l'ouest. Certaines données montrent une migration de l'ouest de la Russie vers le reste de l'Europe.
Écrit par : Thomas FERRIER | 13/07/2014
Écrit par : Anton Cusa | 14/07/2014
Écrit par : Anton Cusa | 14/07/2014
Quant à l'intérêt de l'article, en effet il est de montrer la relative homogénéité entre Européens et l'absence de subdivisions pertinentes en leur sein.
Écrit par : Thomas FERRIER | 14/07/2014
Écrit par : Anton Cusa | 14/07/2014
Les commentaires sont fermés.