
L'Europe est un tout unique
01/03/2010
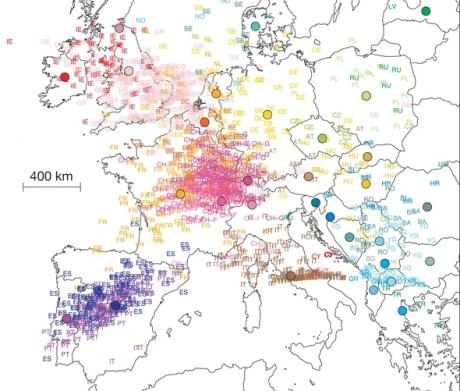
| GENETIQUE – Le profil génétique d’un Européen permet de savoir de quelle région il vient… En faisant analyser vos gènes, on pourra bientôt savoir si vos aïeux venaient de Picardie, de Bavière ou de Lituanie. C’est en tout cas ce que promettent les résultats de deux récentes études de génétique des populations en utilisant une nouvelle technique d’exploration du génome humain permise par les puces ADN : l’analyse des mutations individuelles (SNP). Des profils et des distances John Novembre et une équipe americano-suisse ont analysé une large base de données répertoriant les 200 000 variations génétiques individuelles d’un échantillon de 1387 Européens vivant dans 36 pays. A l’aide d’outils statistiques, ils ont ensuite établi un profil génétique numérique pour chacun des sujets, ce qui leur a permis de déduire une distance génétique entre chaque sujet. Géopolitique des mutations Ils se sont ainsi aperçus que la distance génétique entre deux individus était à peu près proportionnelle à la distance entre les régions où ils vivaient. En projetant graphiquement ce résultat, ils ont ainsi constaté que la carte des profils génétiques recouvrait la carte géopolitique de l’Europe. Ces travaux ont été publiés dimanche dans la revue « Nature » ; les résultats rejoignent d’ailleurs ceux déjà obtenus dans une étude similaire publiée début août dans la revue « Current Biology » et dirigée par le généticien hollandais Manfred Kayser. Généalogie génétique Les deux études montrent qu’en dépit des échanges incessants, des migrations et des invasions, les Européens gardent une odeur de terroir dans leurs gènes. « Cela rend envisageable la mise au point de tests génétiques permettant de connaître les origines géographiques d’un individu » explique John Novembre à 20Minutes.fr « sur notre carte géo- génétique, d’après leur profil, 50% des individus se retrouvaient à moins de 310 km du centre de leur pays d’origine, et 90% à moins de 700 km ». Pas de races européennes Toutefois, le chercheur tient à préciser que cela n’implique pas l’existence de « sous-races » européennes génétiquement différenciées. « En moyenne, les différences génétiques entre populations européennes sont négligeables : sur une échelle de 0 à 1 (1 représentant deux génomes complètement différents) le degré de différentiation génétique est d’à peine 0,004 entre régions européennes ; absolument pas de quoi parler de sous-races, en tous cas au sens biologique » poursuit John Novembre « la possibilité de déterminer la population d’origine de chaque sujet n’a ici été possible qu’en prenant en considération les dizaines de milliers des mutations ponctuelles dont il était porteur ; on a ensuite repéré celles qui étaient les plus fréquentes dans les différentes populations européennes pour inférer l’origine géographique de chaque individu ». Yaroslav Pigenet |
25 commentaires
Correction : Il demeure que 4 des 6 principaux haplogroupes rencontrés chez les Européens se retrouvent aussi bien en Asie qu'en Afrique du nord, ainsi qu'au Proche et Moyen-Orient. Les haplogroupes E1b1b et J sont, en effet, caractéristiques de toutes les populations des deux rives du pourtour méditerranéen et leur présence sur la rive nord est mis en rapport avec la néothilisation de l'Europe par des migrations massives en provenance du Proche-Orient. Les haplogroupes R1a et R1b sont mis en rapport avec l'expansion indo-européenne et se retrouvent, aujourd'hui encore, non seulement chez les populations asiatiques de langue indo-européenne, mais également chez les très nombreux peuples turcs qui, de l'Oural à l’Altaï, se sont massivement mélangés aux Indo-Européens à travers l'histoire. Il est non moins intéressant de mentionner qu'un sous-groupe de l'haplogroupe R1b, que l'on associe d'un point de vue ethnolinguistique aux Indo-Européens occidentaux, se retrouve massivement au sein de populations sub-sahariennes purement négroïdes (jusqu'à 95 % dans des lignages masculins de tribus Bantous du nord du Cameroun). Cela atteste, semble-t-il, que les lignages d'haplogoupes sont beaucoup plus anciens que les processus de "raciation" qui ont aboutit à l'apparition des grandes races humaines actuelles (et infiniment plus anciens aussi que les processus d'ethnogenèse et de glottogenèse, qui relèvent, eux, plutôt de l'évolution sociale des populations). Les plus anciens ancêtres d'un pur négroïde peuvent être en effet plus proches des plus anciens ancêtres d'un pur europoïde que des plus anciens ancêtres d'un autre pur négroïde. Le fait qu'il soit possible de caractériser génétiquement aujourd'hui des grands ensembles de populations humaines correspondant "grosso modo" aux grandes races humaines (I, R, J, E1b1b pour les europoïdes, E1b1a, E2, A, B pour les négroïdes, D, Q, C,O pour les mongoloïdes, etc.) ne doit pas induire en erreur. La "raciation" est un phénomène plus tardif, qui s'est produit au sein de populations que leurs migrations avaient simplement conduites sur des mêmes territoires. Les correspondances actuelles entre grandes races et haplogroupes s'expliquent par le fait que les porteurs de chaque haplogoupe (ou de chaque sous-groupe) ne migraient pas individuellement, mais le plus souvent collectivement, en s’installant avec l'essentiel de ses effectifs sur un même territoire (sans que chaque membre se disperse aux quatre coins du monde). Bien que les porteurs de R, de J, les I et de E1b1b eussent des ancêtres différents, le fait qu'ils se fussent retrouvés sur un vaste territoire commun (où les avaient portés leurs migrations) les a conduit à avoir une longue évolution biologique commune ayant déterminé l'apparition de caractéristiques morphologiques communes (conséquence de l'isolement, de la sélection, du climat, etc.). Le fait que les principaux porteurs de R1b se fussent dirigés vers le nord de Caucase et se fussent installés dans les régions pontico-ouralo-caspiennes, où leur évolution ethnolinguistique et culturelle les conduira à former beaucoup plus tard les Indo-Européens (beaucoup plus tardivement d'ailleurs que leur "raciation" en europoïdes, commune à bien d'autres populations), n'a pas empêché dans ce cas d'autres porteurs de cet haplogroupe de migrer précocement en Afrique centrale, où ils auraient à connaitre des processus de "raciation", d''ethnogenèse et de glottogenèse complètement différents. Le fait d'avoir en commun ses premiers ancêtres, non pas avec les représentants des souches raciales, ethniques, linguistiques et macro-culurelles auxquelles ont appartient, mais avec des représentants de souches complètement différentes des siennes, ne signifie donc pas grand chose en terme d'identité.
Sans oublier bien-sûr la présence massive de l'haplogroupe spécifique (N) des populations mongoloïdes altaïques et ouraliennes du nord-est de l'Asie (tels que les Yakoutes) parmi les Finlandais, les Baltes ou les Grands Russiens...
l'haplogroupe N (N1c et N1b) est d'ailleurs l'un de très rares à être complètement "trans-racial", les représentants de ce lignage devant être répandus du détroit de Béring à l'est de la Scandinavie bien avant leur "raciation" en mongoloïdes et en europoïdes. C'est sans doute la seule explication pour justifier sa présence tout aussi massive au sein de populations radicalement différentes à un point de vue "racial".
Il faut se méfier de ces données qui peuvent parfaitement remonter à une époque bien plus ancienne, par exemple au stade erectus (comme pour les groupes sanguins), surtout si on admet une sapientisation progressive dans des niches spécifiques.
Quant à l'origine que vous donnez des IE, elle n'est pas compatible avec les données linguistiques, les seuls fiables en la matière, ni avec la mythologie comparée (et surtout avec sa symbolique associée).
Au-delà de la subdivision au sein de l'espèce en quatre sous-ensembles puis des adaptations in situ au sein de chaque ensemble, on ne peut rien déduire.
Vous n'avez pas tort sur le processus initial mais sur la chronologie. Ce changement d'échelle change tout.
Enfin, vous passez sous silence la forte homogénéité des populations européennes que cet article évoque, en dépit des quelques haplotypes "transphyliques" que vous évoquez remontant sans doute à une époque bien plus ancienne (probablement avant qu'homo erectus ne se divise en branches).
Cordialement.
En clair, les gènes aussi ont une histoire. Certains de nos gènes remontent même à l'époque des eucaryotes. Que certains se retrouvent un peu partout ou de manière "baroque" ne change pas le fait qu'on en partage beaucoup plus en commun entre européens qu'avec quiconque.
Correction : Je ne repasserai évidement pas en revue la série impressionnante et variée de "données" qui conduit la grande majorité des spécialistes à localiser le terreau ethnoculturel et ethnolinguistique initial des proto-IE dans cette région médiane entre l'Europe centrale continentale et l'Asie qu'est la zone pontico-caspienne (ce n'était pas d'ailleurs le sujet). Je ne sais pas très bien à quel patrimoine génétique spécifique seulement aux Européens actuels vous vous referez, mais il ne me semble pas dépourvu d'intérêt de faire observer que la distribution des seuls gènes pouvant servir à l'heure actuelle de marqueurs généalogiques fiables des populations (les haplogroupes) atteste clairement que les seules composantes génétiques de ce type spécifiques seulement aux Européens actuels sont "grosso modo" les groupes I1 et I2 (héritées très probablement des premiers hommes modernes d'Europe, entendez les hommes de Cro-Magnon). Le reste des principales composantes génétiques (E1b1b, J2, R1b, R1a et N) sont massivement communes aux Européens, aux Nord-Africains, au Proche et Moyen-Orientaux, aux populations asiatiques de langue indo-européenne, aux peuples Turcs (qui sont souvent des IE turquisés), ainsi qu'aux populations du Grand Nord asiatique. Autrement dit, beaucoup d'Européens peuvent être génétiquement plus proches d'autres populations qu'ils ne le sont d'autres Européens. Et pour cause, la réalité européenne n'a évidement pas la même profondeur chronologique que les souches anthropobiologiques dont il est question ici...Je ne comprends très bien votre volonté de faire remonter la présence et la diffusion de ces haplogroupes à l'époque de l'homo Erectus alors que leur présence et leur diffusion est plus ou moins clairement mise en rapport avec des faits beaucoup plus récents et assez bien connus : peuplement des hommes de Cro-Magnon en Europe (I), processus de néolithisation de l'Europe par des migrations massives originaires depuis le Croissant fertile (E1b1b, J), indoeuropéanisation de l'Europe et d'une partie de l'Asie (R1a, R1b), ancienne matrice paléo-asiatique commune de la Scandinavie orientale au détroit du Béring (N), etc. La présence massive des groupes E (E1b1a, le groupe majoritaire chez les subsahariens) et R1b (R1b-V88) chez des purs négroïdes, ainsi que la présence généralisée du groupe N, tant chez des purs europoïdes que chez des purs mongoloïdes (de la Scandinavie au détroit du Béring) indique semble t-il que l'apparition (raciation) des caractéristiques somatiques, anatomiques, phénotypiques, morphologiques et physiologiques qui subdivisent l'homme moderne en grandes "races" est un phénomène assez récent, ne pouvant pas servir de justification à la théorie de la polygenèse de l'homme moderne. Car on demeurait bien en peine d'expliquer alors que les plus anciens ancêtres d'un pur europoïde Breton porteur de R1b soit plus liés aux premiers ancêtres d'un pur négroïde Bantou porteur du même groupe qu'aux premiers ancêtres d'un pur europoïde porteur d'un autre groupe. Je me garderai pourtant de sacrifier à la correction politique, en rappelant encore une fois qu'il est parfaitement possible de caractériser, sur la base des haplogroupes, des grands ensembles de populations humaines correspondant "grosso modo", pour des raisons déjà évoquées, aux grandes races actuelles : c'est que les porteurs de ces haplogroupes formaient le plus souvent, déjà avant leur "raciation", des groupes de populations endogames migrant et s'installant sur un territoire avec l'essentiel de leurs effectifs. Les haplogroupes ont une histoire, mais ils sont loin de se retrouver un peu partout d'une manière "baroque" comme vous le prétendez, non sans quelque malice, pour réfuter complètement leur valeur de marqueur généalogique.
erratum : "originaires du Croissant fertile", et non pas "depuis le Croissant fertile"
Pour information, le dernier ancêtre commun à tous les sous-groupes de l'haplogroupe R1b (stock à partir duquel se formeront notamment les proto-IE) est daté d'environ 16000 ans, on est donc très loin de l'homo Erectus (bien qu'il fût manifestement antérieur aux processus de "raciations" au sein des Homo sapiens sapiens, il est bien postérieur au processus de "spéciation" qui a conduit à l'apparition de ces derniers). Quant au plus récent ancêtre patrilinéaire commun à tous les hommes modernes (haplogroupe A du chromosome Y), il daterait de 140000 ans. Je ne sais où vous êtes allés chercher que les gènes actuelles de l'homme anatomiquement moderne à valeur de marqueurs généalogiques (haplogroupes de l'ADN chromosomique Y ainsi que ceux de l'ADN mitochondrial) dateraient de l'époque de l'Homo Erectus. Je me répète, mais l'idée, très minoritaire, d'une évolution indépendante de l'Homo sapiens sapiens à partir de l'Homo Erectus en plusieurs endroits différents, ainsi qu'en des époques bien distinctes (polygénisme), ne sauraient se baser sur les caractéristiques morphologiques, somatiques ou phénotypiques qui subdivisent l'homme moderne en grandes races. Car il serait difficile alors d'expliquer que les derniers ancêtres communs de populations très différentes "racialement" (Bretons et Bantous massivement de groupe R1b, ou Finlandais, Estoniens, Lithuaniens, Samoyèdes ou Yakoutes massivement de groupe N, etc.) pussent remonter à une époque aussi basse, tandis que des derniers ancêtres communs que ces populations ont avec d'autres populations apparentées "racialement" remontent à une époque beaucoup plus haute.
Pour R1b, voir notamment : "Ancient History of the Arbins, Bearers of Haplogroup R1b, from Central Asia to Europe, 16,000 to 1500 Years before Present, de Anatole A. Klyosov"
http://www.scirp.org/journal/PaperInformation.aspx?paperID=19567#.U8FEApg0uIg
erratum : "ne saurait", et non pas "ne sauraient" (...l'idée, très minoritaire, d'une évolution indépendante...ne saurait...)
La datation des données génétiques reposant sur les haplogroupes est contestée. L'évolution morphotypale au sein de l'humanité est de toute évidence beaucoup plus ancienne. Il est impossible que l'homo erectus en Asie et l'homo sapiens en Asie aient évolué de manière très similaire en étant le fruit de deux vagues migratoires venues d'Afrique séparées de plus d'un million d'année.
Que les Néanderthaliens et les hommes européens modernes présentent des caractéristiques morphotypales extrêmement proches est également une donnée importante. Même si les uns ne descendent pas des autres, mais sont deux évolutions de l'homo antecessor, l'évolution proprement européenne d'homo erectus.
La théorie de l'évolution multirégionale, qui n'interdit pas des flux géniques à la marge (pouvant largement expliquer ce que vous évoquez), repose avant tout sur des données anthropologiques et archéologiques.
Considérer que l'ancêtre commun aurait 140.000 ans et viendrait d'Afrique est une théorie, fortement idéologisée, reposant sur une datation basse des données génétiques. Or, si ancêtre commun incontestable il y a, c'est homo erectus.
Le plus logique est donc une évolution in situ expliquant aisément l'extrêmement forte homogénéité de la population européenne, n'interdisant pas à la marge l'introduction d'exo-gènes. En clair, un départ d'homo erectus, passage par le Caucase (homme de Dminissi), implantation dans l'ouest de l'Europe (antecessor), évolution en deux sous-ensembles, qui deviendront les néanderthaliens, hyper-adaptés à l'environnement glaciaire, inadaptés à tout autre environnement, et les proto-europoïdes, installés dans l'extrême sud-est de l'Europe, et qui peupleront ensuite le reste de l'Europe et une partie de l'Asie, puis le nord de l'Afrique.
On pourrait d'ailleurs grossir le raisonnement d'exemples en invoquant le groupe E (que l'ont date de 55000 ans). Les derniers ancêtres communs d'un europoïde kosovar (ou d'un europoïde Grec, etc.) du sous-groupe E1b1b et ceux d'un négroïde subsaharien du sous-groupe E1b1a (le groupe majoritaire des négroïdes) remontent à une époque beaucoup plus basse que l'époque à laquelle remontent leurs derniers ancêtres communs avec toutes les autres populations humaines.
Comme je vous l'ai dit, je conteste deux choses:
- la confusion entre flux génique et ascendance commune
(la notion d' "ancêtres communs" est-elle même un présupposé d'adhésion à la théorie monophylétiste)
- une datation qui repose là aussi sur le même présupposé
Qu'un flux génique ait pu introduire des gènes qu'on retrouve prioritairement ailleurs au sein de la population européenne, sans que cela ne remette en cause son homogénéité (nécessairement relative), personne ne le conteste.
Enfin, il faut prendre en compte l'ensemble des gènes, ce que cette étude fait, et celle-ci implique une évidence, l'origine commune de toutes les populations europoïdes.
La linguistique là encore permettant de déterminer une origine commune à toutes les langues des populations europoïdes, amenant à saisir l'existence d'une macro-famille préhistorique qu'on peut appeler "nostratique" ou plutôt "europique", et comprenant le proto-indo-européen, le proto-ouralien, le proto-afrasien, le proto-kartvélien... mais aussi le basque, le sumérien, les langues indusiennes... etc
N'étant pas biologiste moléculaire, je me garderai bien de porter des jugements personnels (comme vous le faites) sur la datation absolue de ces gènes à valeur de marqueur généalogique. Je me contente simplement de rappeler les théories accréditées par les spécialistes, dont je ne m'étonne guère que vous en contestiez la valeur, car c'est ainsi que vous procédez toujours lorsque les résultats scientifiques ne sont pas conformes à votre praxis identitaire. Mes observations à caractère plus personnel se bornaient juste à vous faire observer que les composantes génétiques à valeur de marqueur généalogique d'un europoïde peuvent être beaucoup plus proches phylogénétiquement des composantes correspondantes d'un négroïde ou d'un mongoloïde que de celles d'un autre europoïde. Le phénomène que j'évoque n'a rien de marginal, car le taux de fréquence de ces haplogroupes communs est considérable (N1b chez 63.2 % contre 80 % chez les Yakoutes, et ce n'est qu'un exemple). Nous avons affaire à des populations qui ont massivement la même hérédité biologique du point de vue des haplogroupes sans que cela correspondît le moins de monde à un autre phénomène, dont le fondement héréditaire est pourtant non moins évident : la race. Difficile de ne pas en inférer que l'un de deux phénomène est plus tardif que l'autre ("raciation" plus tradive que le lignage d'haplogroupe). Ce type de proximité génétique et phylogénétique (dans les arbres de parenté) plus forte entre un europoïde et un non-europoïde qu'entre deux europoïdes devrait pourtant vous interpeler quelque peu...Ce phénomène de proximité génétique incontestable, ainsi que la parfaite et totale interfécondité de toutes les grandes races humaines actuelles, amèneraient à réfléchir tout esprit honnête et sérieux. Mais il semble que vous préfériez, pour votre part, vous plaire à imaginer je ne sais quelle proximité génétique supérieure avec les membres d'une autre espèce : l'Homo neanderthalensis. Au cas où vous ne l'auriez pas tout à fait compris, cela revient à sauter un rang taxinomique, celui de l'espèce. Dans un autre ordre d'idée, les concentrations beaucoup plus importantes sur la rive sud de la Méditerranée des stocks d'haplogroupes à partir desquelles se produira la "raciation" des protoméditerranéens (E1b1b, J1 et J2), ainsi que la néothilisation plus précoce du Proche et Moyen-Orient, attestent aussi, ne vous en déplaise, que l'origine des europoïdes ne doit pas être recherchée uniquement dans le Sud-Est de l'Europe. Je me demande souvent où vous allez chercher certaines de ces théories pour le moins fixistes. Indifféremment de toutes ces considérations, il appert de tout cela que votre énoncé initial, qui entendait fonder l'unicité des Européens actuels sur la génétique des populations, est bien sujet à révision.
Je réponds à votre dernier commentaire. Relisez-moi donc attentivement, je n'ai pas contesté le moins du monde que les arbres de parenté obtenus à partir du croisement des données anthropobiologiques, linguistiques, voire macro-culturelles, révélaient des corrélations et des apparentements importants entre lignages d'haplogroupes, souches macro-raciales, souches (macro)linguistiques et souches macro-culturelles. J'ai juste fait observer que toutes ces choses ne s'agglutinent pas d'une manière aussi mécanique et déterminée que l'ont pourrait le croire au premier abord. Il faut juste se méfier des illusions rétrospectives de la nécessité. Les porteurs des haplogroupes I, J, R et E1b1b n'étaient tout à fait obligés dans le fond de migrer et de s'installer collectivement, avec l'essentiel de leurs effectifs, entre l'Atlantique et l'Oural (et entre l'Afrique du nord et le Moyen-Orient) pour connaitre une "raciation" commune en europoïdes, y développer des langues (indo-européennes, caucasiques, afro-asiatiques, asianiques, etc.) appartenant sans doute à même ensemble macro-linguitique, ou des civilisations plus ou moins parentes.
erratum : Le phénomène que j'évoque n'a rien de marginal, car le taux de fréquence de ces haplogroupes communs est considérable (N1b : 63.2% chez LES FINLANDAIS contre 80 % chez les Yakoutes, et ce n'est qu'un exemple). J'avais oublier le nom de FINLANDAIS
Il faut se méfier de la génétique, qu'on croit souvent plus avancée qu'elle ne l'est. Le tableau des allèles de Cavalli-Sforza converge en outre littéralement avec les données de l'anthropologie physique par exemple. Attention à ne pas surévaluer une donnée dont l'origine n'est pas forcément si évidente que ça.
L'espèce humaine n'est pas différente des autres espèces mammifères, à savoir une subdivision ancienne en plusieurs sous-espèces. Ce qui implique une origine commune à tous les membres d'une même sous-espèce. Quant à l'interfécondité, elle est établie entre le lion d'Afrique et le lion d'Asie, par exemple. Ce qui ne signifie pas une différenciation récente.
J'ai affirmé que l'homo sapiens europaeus descendait comme l'homo neanderthalis d'un ancêtre commun. Je n'ai pas affirmé interfécondité entre les deux. Je pense plutôt à une variante "aberrante" hyper-spécialisée de l'un dans un environnement spécifique. En clair, les néanderthaliens se seraient rapidement isolés et différenciés de l'autre branche cousine.
La seule explication reste un flux génique constant, pas forcément important néanmoins, ce qui est admis par les partisans de la théorie de l'évolution multirégionale.
Quant à l'interprétation des données, qui est au moins aussi importante que les données elles-mêmes, elle repose souvent sur des "postulats de principe", comme dans la fameuse affaire Clovis, où les chercheurs refusaient de fouiller en dessous de -12.000 ans, alors qu'on a fini par trouver la preuve d'un peuplement plus ancien de l'Amérique (sans doute solutréen).
Anthropologues et généticiens "médiatiques" s'opposent. J'ai évoqué mes propres intuitions avec des chercheurs en ce domaine, qui ont reconnu la faiblesse du discours sur les fameux "haplotypes" et aussi sur leur caractère de marqueur d'ascendance.
Quant à savoir si cela est dominant, visiblement c'est moins de 0,004% en moyenne.
Cet article donne de l'importance à la notion d'homogénéité donc de profonde parenté, ce qui ne signifie pas uniformité. Il n'est en rien remis en cause.
Je précise que la génétique n'est pas ma spécialité, et je ne prétends pas qu'elle le soit, mais que je suis très critique sur une lecture qui en est faite et qui ne correspond pas à des faits de base. Je note toutefois que vous ne niez pas l'existence de sous-espèces au sein de l'espèce humaine. Cela contredit de fait une partie de votre argumentaire.
Je n'ai ceci dit pas besoin de m'appuyer sur des données génétiques. Je trouve simplement que cet article était intéressant car rappelant une évidence.
Je confesse avoir énormément de mal à vous suivre, Thomas. Contrairement aux données génétiques obscures, à la distribution aléatoire (où la variabilité intraraciale peut être bien supérieure à la variabilité interraciale), sur la base desquels vont affirmiez pouvoir démontrer, semble-t-il, l'unicité et l'ascendance biologique commune des Européens actuels, la valeur phylogénétique (mais non systématique) de marqueur d'ascendance des particularités du génome humain que représentent le chromosome Y et l'ADN mt est en revanche très largement acceptée par les spécialistes, et ce d'autant plus que ces données correspondent le plus souvent (mais pas systématiquement) non seulement aux connaissances phylogénétiques des classifications linguistiques, mais également aux grandes races humaines. La génétique se révélerait donc éminemment secourable selon vous quand cela vous arrange (lors même que les données fussent alors également, ou aléatoirement, répartis dans toutes les populations humaines), pour devenir aussitôt suspecte lorsque cela ne sied plus à votre praxis, quand bien même elle mettrait alors au jour des marqueurs plus spécifiques et très inégalement distribués entre les groupes raciaux et ethnolinguistiques. Votre hypercriticisme est véritablement acrobatique ! Le génome humain n'étant pas entièrement exploré, le terrain n'est assurément pas aussi ferme qu'en matière de démonstration mathématique ou de logique apodictique, mais je invite néanmoins à observer de nouveau les quelques faits simples que je vous ai rappelé : distribution massive des R1a et R1b dans toutes les régions, asiatiques et européennes, indoeuropéanisées ainsi que parmi les peuples turcs répartis entre l'Oural et l'Altaï (et d'ailleurs), où les Turcs se sont intimement mélangés aux Iraniens orientaux cavaliers archers nomades des steppes. Présence importante de J-1, J-2 et de R1b1b sur les deux rives du pourtour de la Méditerranée, et massivement sur la rive sud, d'où provenaient les formations de migrants qui ont colonisé et néolithisé l'Europe. Distribution massive des groupes N1b et N1c chez les populations europoïdes et mongoloïdes, européennes et asiatiques, réparties, depuis des temps immémoriaux, sur presque tout l'ensemble de l'Eurasie septentrionale, de la Scandinavie orientale au détroit du Béring, ce qui correspond très bien à l'ancienne matrice paléoasiatique commune, d'où sont issus notamment les anciens Ouraliens, etc. Je préfère ne pas insister. Ce n'est pas tant l'interfécondité que la proximité génétique dont je vous ai entretenu dans mes messages antérieurs (grande absence de variabilité interaciale pour des marqueurs spécifiques pourtant très inégalement distribués entre les groupes raciaux en règle générale) qui suggère le caractère tardif de la "raciation" chez les hommes modernes. Je vous invite à me lire honnêtement. S'agissant de la notion de race appliquée à l'humanité, il faut pourtant insister sur la caractère assez flou de ce dernier rang taxinomique de la classification du vivant (après celui de l'espèce), à savoir la race, nommée aussi "variété", "forme" ou "sous-espèce". Le caractère flou, et moins insécable, de la race (par rapport aux autres rangs taxinomiques) résulte bien sûr de l'interfécondité des races qui subdivisent et composent une même espèce. Il est évident aussi que, contrairement aux races animales d’élevage domestique, qui résultent d'un isolement et d'une homogénéisation artificielle, le processus de "raciation" chez les hommes modernes est fortement marqué par les multiples mouvements et brassages qu'implique l'histoire sociale et culturelle de ceux-ci (et c'est là bien-sûr une particularité de l'Homo sapiens sapiens). La notion de race n'est pas si pertinente pour cette espèce en ce sens qu'il n'existe jamais de race spécifique à une seule population concrète élémentaire (c'est à dire à elle seule). Il s'en faut de beaucoup par exemple pour qu'une des subdivisions de la grande race europoïde (alpine, nordique, est-baltique, dinarique, méditerranéenne, pamrienne, etc.) fût spécifique à un groupement socio-humain concret et à lui seul (même lorsqu'il s'agit du groupement socio-humain le plus élémentaire, entendez l'ethnie). Les subdivisions concrètes fondamentales et élémentaires de l'humanité résultent avant tout de l'évolution culturelle et sociale des hommes, au moins depuis la révolution du Néolithique (qui a réduit les facteurs d'isolement géographique). Il n'y a pas de groupement socio-humain concret pouvant être nommée alpin, dinarique ou nordique, et ce bien que les processus d'éthnogenèse se produisent au sein de populations qui combinent des stocks génétiques précis. Les découpages très abstraits de la zoologie humaine ne conduisent certainement pas aux découpages concrets de l'ethnologie, de la sociologie et de l'histoire (les populations humaines n'évoluent plus dans un pur état de nature). La notion de grandes races, qui subdivisent plus clairement l'espèce en trois ou quatre grands groupes, est pertinente surtout pour des raisons d'isolement géographique et d'ordre ethno-culturel. Mais là où l'isolement était plus limité, c'est à dire dans les régions frontalières entre grandes races, les mélanges, les mouvements et les interconnexions ont été si réels qu'il est loisible de qualifier ces vastes régions de "zones grises". Il en va ainsi des versants du Sahara ou de la région située entre l'Oural et l'Altaï (etc.).
Je partage globalement votre propos sur les sept dernières lignes de votre texte.
Quant au peuplement néolithique de l'Europe via le moyen-orient pré-afrasien, c'est une hypothèse effectivement majoritaire et assez crédible, mais il ne faudrait pas négliger l'idée que le moyen-orient a pu préalablement être peuplé de populations venues d'Europe, dont les ancêtres des Sumériens et des Hourrites.
Un certain nombre de données laisse penser que le peuplement de l'Europe par l'homme moderne se soit fait de l'est vers l'ouest, mais pas du sud-est vers l'ouest. Certaines données montrent une migration de l'ouest de la Russie vers le reste de l'Europe.
Je pense que vous devriez plutôt insister sur le caractère bien plus limité, par rapport à l'Afrique ou à l'Asie, de la variabilité des marqueurs génétiques d'ascendance (et également de la variabilité raciale) intra-européenne, car vous seriez là sur un terrain infiniment plus ferme que celui du polygénisme. En refusant de plaquer mécaniquement le concept zoologique et abstrait de race sur celui de l'ethnie (le groupement socio-humain concret le plus élémentaire), j'apporte en quelque sorte de l'eau à votre moulin. Car si la variabilité raciale intra-ethnique peut être supérieure à la variabilité raciale inter-éthnique à l'intérieure d'une grande race humaine géographique, cela signifie par exemple que les peuples européens ne sauraient sérieusement de distinguer entre eux sur des critères raciaux (alpins contre dinariques, etc.). Les alpins ou les dinariques n'ont pu former des groupements socio-humains concrets qu'à l'époque du Paléolithique, où les populations pouvaient encore être issues de souches particulièrement limitées en raison de leur isolement géographique. La révolution du Néolithique a en revanche pleinement installé l'espèce dans la dimension sociale et culturelle de son évolution et de son histoire (qui implique des brassages innombrables et inextricables entre subdivisions raciales sur une aire géographique vaste). J'en reviens au polygénisme. Il bien est curieux de vous voir ainsi affirmer que l'apparition des mêmes caractéristiques somatiques, morphologiques, anatomiques, génétiques, physiologiques et phénotypiques aussi différentes des précédentes (spéciation en Homo sapiens sapiens) pussent s'être produites spontanément et indépendamment sur de nombreux points du globe (sans que le dernier ancêtre commun remontât à l'Homo sapiens sapiens donc) en admettant, tout à la fois, la monogenèse et l'ascendance commune à l'intérieure des grandes races qui subdivisent ce même Homo sapiens sapiens. La raciation en europoïdes impliquerait nécessairement la plus haute ascendance commune (malgré le démenti de la génétique), tandis que la spéciation en Homo sapiens sapiens n'impliquerait point, quant à elle, la monogenèse...Je me répète, mais la variabilité intra-raciale parfois supérieure de certains marqueurs génétiques d'ascendance (N, E, R) semble bien indiquer que la raciation est un phénomène assez tardif, produit davantage sous l'effet du climat, de l'écologie ou de la thermorégulation. Dans le même ordre d'idée, le hiatus concernant les marqueurs génétiques d'ascendance entre les négro-africains et les représentants des races négritos et mélano-indiennes (d'Asie), également qualifiées de négroïdes, est pourtant ainsi relativisé par de nombreux marqueurs polymorphiques communs, tel le prognathisme etc., semble bien confirmer ce raisonnement. Pour ce qui est des mouvements nord/sud dans le pourtour méditerranéen, la seule "descente" de "proto-européens" vers le sud ne pourrait être attribuable à ma connaissance qu'aux cromagnoïdes (mechtoïdes) paléolithiques de l'actuel Maghreb, largement absorbés en suite par les proto-méditeranéens. Il semble que les traces les plus nettes de ces cromagnoïdes se retrouvent chez les Kabyles ou chez les anciens Guanches.
erratum : "...Dans le même ordre d'idée, le hiatus concernant les marqueurs génétiques d'ascendance OBSERVABLE entre les négro-africains et les représentants des races négritos et mélano-indiennes (d'Asie), également qualifiées de négroïdes, est pourtant ainsi relativisé par de nombreux marqueurs polymorphiques communs, tel le prognathisme etc. CELA semble bien confirmer ce raisonnement...."
Les populations mélano-indiennes ou "dravidiennes" ne sont pas considérées comme afroïdes.
Quant à l'intérêt de l'article, en effet il est de montrer la relative homogénéité entre Européens et l'absence de subdivisions pertinentes en leur sein.
Il est vrai que la classification raciale des mélano-indiens fait encore débat (malgré certaines similitudes polymorphiques avec les négroïdes ). Le cas de la race négrito, à laquelle on peut adjoindre d'ailleurs ladite race mélanésienne (Papous, Kanaks, etc.), est pourtant loin d'être mal à propos en la circonstance. Nous avons bien là des populations qui présentent des similitudes polymorphiques frappantes avec les négro-africains tout en ayant des marqueurs génétiques d'ascendance qui les en éloignent beaucoup phylogénétiquement. J'apporte une petite correction à cette phrase précédente, dont la première partie est quelque peu obscure : "Je me répète, mais la variabilité intra-raciale parfois supérieure de certains marqueurs génétiques d'ascendance (N, E, R) semble bien indiquer que la raciation est un phénomène assez tardif, produit davantage sous l'effet du climat, de l'écologie ou de la thermorégulation." Je voulais bien-sûr dire par là que des masses de populations appartenant à une même grande race humaine peuvent être phylogénétiquement plus proches (derniers ancêtres communs remontant à une époque plus basse dans l'arbre de parenté génétique) de populations appartenant à une autre grande race humaine que d'autres populations appartenant au même ensemble racial qu'elles (Finlandais europoïdes plus proches phylogénétiquement des Yakoutes mongoloïdes, les deux étant porteurs du marqueur N1c). Dans ce cas, bien qu'il s'agisse de marqueurs génétiques d'ascendance, la variabilité intra-raciale est supérieure à la variabilité inter-raciale.
Les commentaires sont fermés.